
At the 2008 ASM General Meeting, the Division U keynote lecture was headed by Tom Gillis of the National Hansen's Disease Program. His talk described the same work I cited in the previous article, which showed the immense amount of pseudogenes in the M. leprae genome.
Gillis was interested in elucidating the role of these pseudogenes. This included asking whether or not these genes are transcribed and translated. If these pseudogenes are not providing any function, then it stems that the cells will not put energy towards their expression.
The work he discussed showed that ~44% of all M. leprae transcription was due to pseudogene expression. There doesn't appear to be a locational bias for pseudogene transcription either. Looking closely at 10 pseudogenes downstream of full-length genes, only 8 produced full-length transcripts.
More indepth in silico analysis shows that all these pseudogenes are unilogs (no duplicates present in the M. leprae genome), the vast majority lack a strong Shine-Delgarno sequence upstream, ~75% lack a translational start codon, and ~98% have one or more in-frame stop codons inserted.
This indicates that a very small percentage of pseudogene transcripts actually create a full-length translational product. So, although the cells still create the transcript, few (if any) resources are put towards creating a functional (or detrimental) protein product.
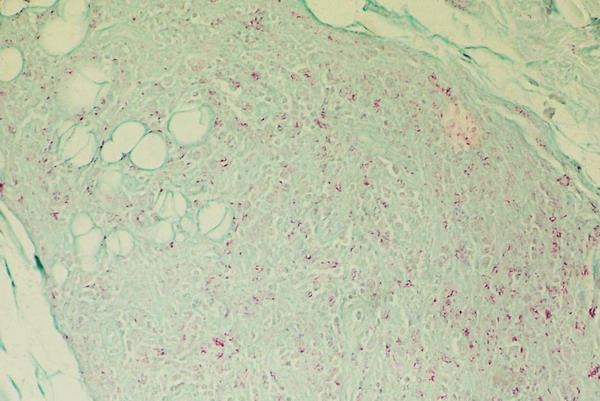
I also picked up some interesting epidemiological facts of the M. leprae genome. For one, the global M. leprae population is nearly clonal (1 polymorphism to 20,000bp compared to 1:5000 for M. tb.). However, variation in SNPs can be seen in local populations. In looking at ~60 cases from a town in India, the bug had a higher rate of diversity than compared to 3 cases in the South Eastern US or to 20 wild armadillos. Furthermore, the US cases and the wild armadillo cases were strikingly similar on an SNP scale.
I think an important point to take home from this is that M. leprae is still an evolving organism, and we are only catching a snapshot in time. It is a prime example of a parasite that has come to depend greatly on its host and has lost the ability to function outside said host.
Other articles of mine that may be of interest
Reductive Evolution in Mycobacterium Leprae
Evolution of Phage Capsid and Genome Size (Another 2008 ASM General Meeting Lecture)
 Viruses come in all shapes and sizes. From the very small, such as the
Viruses come in all shapes and sizes. From the very small, such as the 
