 Produce borne diseases have recently been gracing the front pages of our media. Our spinach has E. coli, our onions have Hepatitis A virus and E. coli, our strawberries have Listeria, and our tomatoes and peanut butter have Salmonella. Not to mention the countless tons of ground beef tainted with pathogenic E. coli.
Produce borne diseases have recently been gracing the front pages of our media. Our spinach has E. coli, our onions have Hepatitis A virus and E. coli, our strawberries have Listeria, and our tomatoes and peanut butter have Salmonella. Not to mention the countless tons of ground beef tainted with pathogenic E. coli.Common sense says that washing and proper handling of our food should simply be enough to prevent illness outbreaks. It has now been hypothesized that many bacteria were able to "hide" within and among the plant cells, protected by their sturdy cell wall. Or even that some pathogenic bacteria were able to enter the cells and remain protected from traditional washing methods.
An article in this month's Applied and Environmental Microbiology looks at a much different method of bacterial survival on produce. They hypothesize that these bacteria are taking refuge in various protozoa, and subsequently are protected from washing and other sanitation methods due to being held either within the cell or an exogenous cell-derived vesicle.
The authors used protozoal isolates from store-bought spinach and romaine lettuce. They measured the concentrations of various microbes from the surface, and cultured a set of naturally occurring protists from these samples.
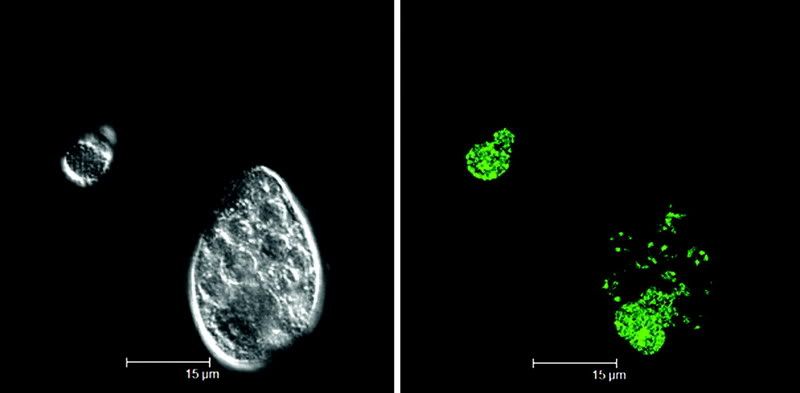
They show that not only are certain pathogenic bacteria (like E. coli 0157:H7) able to be taken up by protozoa, but that in many cases, they survive to be ejected into the world around, surrounded by a vesicle. The flourescent image at above shows this event happening with E. coli 0157:H7 and a strain of Tetrahymena.
Previous studies have shown that the bacteria in these extracellular vesicles are able to survive for the long-term, even in harsh conditions.
This paper shows us that microbial ecology is much more complex than we perceive, and that simple and obvious solutions may not always be the best. We now know that wild protozoa living on our produce can sequester pathogenic bacteria, and furthermore wet produce allows vesicles to be created containing viable bacteria.
This makes me wonder about the current trend in grocery stores to spray there fresh produce with water at specific time intervals. If protists are doing what this study shows they are doing, and vesicles form on wet leaf surfaces, this practice may be something we must reconsider for our food safety.
Gourabathini, P., Brandl, M.T., Redding, K.S., Gunderson, J.H., Berk, S.G. (2008). Interactions between Food-Borne Pathogens and Protozoa Isolated from Lettuce and Spinach. Applied and Environmental Microbiology, 74(8), 2518-2525. DOI: 10.1128/AEM.02709-07



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}